薬物治療の標的分子を解明し、病気の治癒やQOLの改善を目指します。
分子薬物治療学研究室
![]()
〒920-1192 金沢市角間町 金沢大学薬学系
研究概要Research
エルゴチオネイン物語
私たちの研究室では、トランスポーター(膜輸送体)の研究している過程で、アミノ酸の一種であるエルゴチオネインが膜輸送体の生体内基質(膜輸送体が体内で運んでいる物質)であることを見出しました。以来10年余りにかけて、膜輸送体・エルゴチオネイン・私たちの身体、の間の関係について研究してきました。ここではその途中経過を、他の研究グループによる成果も一部交えて、ご紹介します。
1.膜輸送体OCTN1欠損マウスの作製とメタボローム解析
~エルゴチオネインは膜輸送体によって食物から体内・細胞内へ取り込まれる~
膜輸送体は、細胞膜に埋め込まれた構造をした膜タンパク質です。身体の中で作られる内因性化合物、あるいは外から取り込む外因性化合物を認識し、それらの細胞内への取り込みや排出を行います(図1)。したがって膜輸送体は、それら物質の細胞の内と外との間の濃度比を規定します(図1)。そのため、栄養物や薬物の身体の中での動き(体内動態)に深く関与するばかりでなく、疾患部位に存在する膜輸送体やその膜輸送体が輸送する化合物を利用すれば、当該部位を標的とした疾患の治療や、病態の変化を追跡するバイオマーカーなどに応用できる可能性があります。
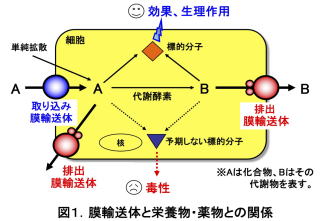
私たちが研究してきた膜輸送体の1つに、carnitine/organic cation transporter 1 (OCTN1)があります。Solute
carrier (SLC) superfamilyという膜輸送体の巨大なfamilyに属するため、SLC22A4という番号がふられています。OCTN1は以前に金沢大学で発見された膜輸送体です。OCTN1を培養細胞に導入すると、さまざまな有機カチオン性化合物を細胞内に取り込むため、OCTN1はorganic
cation transporterとして位置付けられていました。しかし、生体内で何を輸送しているかは不明でした。一方で、ヒトOCTN1遺伝子は、リューマチ性関節炎1)、クローン病※
(503L (WT) < 503F)2)、自己免疫性甲状腺疾患3)、劣性非症候性難聴 DFNB604)など、さまざまな疾患と関係していることから、生体の恒常性を維持する何らかの重要な物質を輸送する可能性が考えられました。
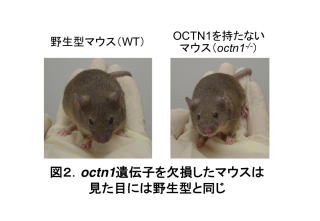
そこで、OCTN1が生体内で何を輸送しているかを解明し、OCTN1の役割を理解するため、octn1遺伝子を欠損したマウス(octn1-/-)を作製しました。しかし、野生型(遺伝子正常)のマウス(WT)と比べoctn1-/-には見た目に違いはなく、成長、繁殖、生化学マーカー等もWTと同等でした(図2)。つまり、OCTN1という膜輸送体がなくても普通に成長していることになります。ということは、OCTN1は生きていく上では必要ないということでしょうか?
そこで、そもそも膜輸送体とは、化合物の細胞内外の濃度比を規定するタンパク質であることを思い出し、WTとoctn1-/-の血液や臓器に含まれる生体内化合物の一斉定量(メタボローム解析)を行うことにしました。両マウスで差のあるものが、OCTN1で輸送されている可能性があります。結果、食物由来のアミノ酸の一種であるエルゴチオネイン(ERGO)が、WTの生体内にあまねく存在し、octn1-/-には存在しないことを見出しました(図3)5)。ERGOとは、100年以上も前からその存在が知られている化合物であり、抗酸化作用(酸化ストレスを抑える働き)があります。真菌類やマイコバクテリアといった下等動物が合成できる一方、ヒトやマウスを含む哺乳類はERGOを合成しません。
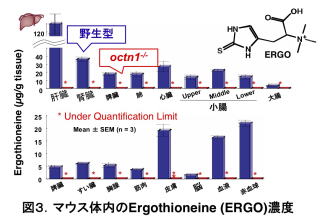
ERGOがOCTN1によって輸送されることは、培養細胞レベルではすでに報告されていました6)。しかし、マウス体内のERGOの濃度を詳細に検討したところ、ERGOを合成しないはずのマウス体内では、調べたあらゆる臓器に数μMから数百μMの濃度でERGOが存在していました(図3)。一方で、octn1-/-ではいずれの血液、臓器にも検出されていません(図3)。このことは、OCTN1がほとんどすべての臓器に発現することと対応し、OCTN1がERGOの体内への摂取と各臓器への分布に働く膜輸送体であることを示しています。ここで、OCTN1遺伝子は魚類、両生類、鳥類、哺乳類など、ヒトやマウス以外のERGOを生合成しない動物でよく保存されています。これらは食物を摂取する動物です。さらにOCTN1は、ヒトやマウス小腸で食物に含まれる成分の血液中への取り込み(吸収)に働く細胞(吸収上皮細胞)の刷子縁膜(血液とは反対側の、食物成分がやってくる側の細胞膜)に局在することが、私たちの研究から分かっています7)。以上のことを考えると、OCTN1は食物由来のERGOを体内に取り込む膜輸送体と考えられます。つまり、体内では合成できないERGOを、わざわざ体内へ取り込むための膜輸送体が、マウスやヒトの体内に存在することを意味します。そしてそれは一体何のためなのか?という新たな疑問が沸き起こります。
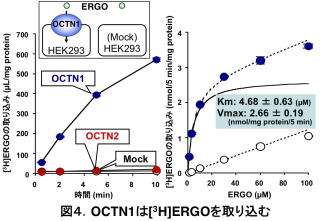
ERGOが体内で何をやっているか、という疑問を解明するには、ERGOが体内でどんな動きをしているか、その体内動態を解明することが重要です。その後の解析から、octn1-/-でERGOがほとんど検出されない理由は、腎臓の近位尿細管というところに存在するOCTN1が、ERGOの再吸収に働いており、octn1-/-ではその再吸収が起こらない(OCTN1がないので)結果、ERGOを摂取しても短時間で尿中へ排泄されてしまうためであることが分かりました5)。OCTN1がERGOを細胞内に取り込む働きのあることも確認されました(図4)5)。また、ERGOを消化管内に投与すると、ほぼ100%が吸収され、octn1-/-では吸収率が低下します7)。つまり、小腸でのERGOの吸収にもOCTN1が働いています。ERGOは極めて水溶性が高く、膜輸送体がないとほとんど細胞膜を透過しません。このことは、HEK293という細胞にOCTN1遺伝子を入れない場合、細胞内にはほとんどERGOが取り込まれていないことからも明らかです(図4)。一方で、小腸からの吸収が良好なのは、OCTN1という膜輸送体があるからであり、OCTN1がERGOを吸収する役割を持っていることを支持しています。小腸で吸収されたERGOは門脈という血管を通って肝臓に入ります。私たちの解析によると、少なくともマウスでは、ERGOのほぼすべてが一度肝臓の細胞内に入ることが分かっています7)。このことは、ERGO濃度が肝臓で、他の臓器に比べ極めて高いことを説明できます。肝臓以外の臓器にもERGOが検出されますが、ごく一部、肝臓を素通りしたもの、あるいは一度、肝臓の細胞内に入ったERGOがやがて肝臓から抜けたものを表しているのではないかと考えています。興味深いことに、ERGOは肝臓の70%を占める肝実質細胞にはほとんど取り込まれまれず、非実質細胞に取り込まれます7)。OCTN1もタンパク質レベルでは肝実質細胞には見られません7)。肝臓はさまざまなホルモンの合成や化合物の代謝、排泄を担いますが、そのほとんどは肝実質細胞の働きです。その意味で、OCTN1とERGOは、肝臓においてかなり特殊と言えます。このことは、後述するERGOの病態における役割と関係します。
2.腸・肝・腎の炎症時でのOCTN1とERGOの役割
~正常な状態ではその働きは見えづらく、病態・炎症に対する防御に働く~
前項で述べたように、OCTN1は炎症性腸疾患の一種であるクローン病やリューマチ性関節炎など、種々の疾患と関連のある遺伝子でコードされるタンパク質です。ERGOは抗酸化物質ですので、細胞内に膜輸送体で取り込まれれば、細胞内で発生する酸化ストレスを抑えることができます。この酸化ストレスは、さまざまな病態時に発生する炎症の際にも見られることから、ERGOがOCTN1によって取り込まれるのは、酸化ストレス、ひいては過剰な炎症を抑えるためではないかと考えることができます(図5)。この仮説が正しければ、OCTN1を持たず、ERGOを体内に摂取することができないoctn1-/-が疾患になれば、野生型(WT)に比べて脆弱であると予想されます。実験で用いるマウスの飼育環境は雑菌などの入りにくい極めて清潔なこと、病気になりやすい高齢まで飼育しないことなどから、octn1-/-が病気に対し脆弱であることが気づかなかった可能性があります。
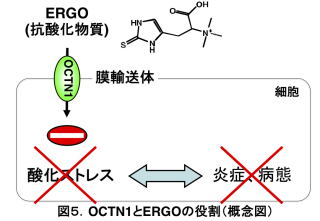
前項で述べたように、経口で摂取されたERGOは、小腸、肝臓を通り、腎臓で再吸収されます。このため、これら臓器の疾患モデルを順にWTとoctn1-/-で作製することにしました。まず、小腸虚血再灌流モデルを作製したところ、WTに比べoctn1-/-では小腸組織の破壊が顕著で、生存率も低いものでした5)。また、デキストラン硫酸を飲水投与することで腸炎モデルを作製したところ、octn1-/-の方が、WTに比べ体重減少や小腸組織の損傷が顕著でした8)。OCTN1がこれら組織損傷に対し防御的に働くことが示唆されます。防御に働いていることは、炎症部位でOCTN1とERGOが高いレベルで存在することによっても裏付けられました。デキストラン硫酸誘発腸炎マウスの小腸刷子縁膜ではOCTN1タンパク質の発現やERGO濃度が、非炎症群に比べ顕著に増加していました8)。クローン病患者の小腸下部でも、対照群に比べた同様なOCTN1発現やERGO濃度の増加が報告されています9)。
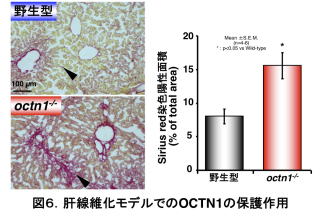
次に、WTとoctn1-/-にジメチルニトロサミンやコンカナバリンAを投与することで肝線維化モデルを作製しました。この場合も、WTに比べoctn1-/-では肝臓での線維化が顕著でした(図6)10)。同時に肝臓での酸化ストレスも観察しており、ストレスマーカーがoctn1-/-の肝臓で顕著なことも確認できました。したがって、OCTN1が肝臓の病態モデルにおいても防御的な役割を果たすことが示唆されました。この時、マウスの肝臓中に含まれるERGO濃度を測定したところ、octn1-/-ではERGOが観察されませんでしたが、WTでは観察され、線維化の進んだ肝臓の方が健康な肝臓よりもERGOが多く含まれていました。ERGOを取り込むOCTN1を観察すると、正常な肝臓では内皮細胞に存在する7)一方、線維化の進んだ肝臓では、活性化された星細胞(伊東細胞)に存在していました10)。星細胞は、普段はビタミンAなどを貯蔵する働きをしますが、活性化されると肝臓の線維化に働くことが知られています。星細胞はOCTN1を発現することでERGOを細胞内に取り込み、線維化を抑えている一方、OCTN1を持たないoctn1-/-の肝臓ではERGOを取り込むことができず、線維化が進んでいたのかもしれません。
慢性腎臓病(CKD)モデルにおいても、WTに比べoctn1-/-で顕著な腎機能の悪化、炎症の亢進、線維化が見られることが示されています11)。この研究では、慢性腎臓病患者においても腎機能低下と血中ERGO濃度が相関すること、腎移植後には腎機能とともにERGOの血中濃度が回復することが示されており、ヒトでの腎臓の病態とOCTN1やERGOとの関係が明らかになりました。
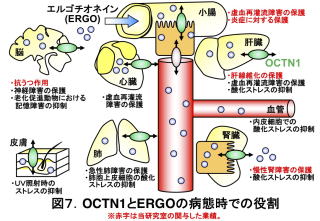
以上、私たちの研究や共同研究で得られた成果を中心に話を進めてきましたが、現在までに、OCTN1とERGOがさまざまな臓器疾患に対し防御的に働くことが示唆されています(図7)。このことは、OCTN1とERGOがほとんどの臓器に存在することと対応します。また、OCTN1とERGOが正常な状態よりもむしろ病態時に重要な役割を果たすと考えれば、快適な環境下で飼育されたoctn1-/-とWTとの比較では、明確な違いが観察されなかったことが説明できるかもしれません。
3.中枢神経系におけるOCTN1の役割
~食物由来の水溶性アミノ酸ERGOが脳へ到達する理由~
ここまで、OCTN1の遺伝子を欠損させたマウス(octn1-/-)を使ってOCTN1やERGOの役割を解明しようと試みてきました。octn1-/-が野生型(WT)と比べて、顕著な違いが観察されなかったのは、両者に違いがなかった訳ではなく、正常な状態で観察していたためにその違いを見逃していたとも言えます。このことは脳におけるOCTN1とERGOの働きを考える上でも注意すべき点かもしれません。マウスはヒトと違って話をすることもできませんし、脳機能の一部に異常があってもそれが見た目には観察されない可能性があります。一方で、食物由来のERGOは脳にも観察されます(図3)。一般に化合物の脳への移行は、血液脳関門と呼ばれるメカニズムによって著しく制限されています。ERGOのような水溶性の高い化合物は、脳に積極的に取り込む膜輸送体が存在しない限り、ほとんど脳には移行しないと考えられています。したがって、
ERGOが脳に存在するということは、脳でも何らかの役割を果たす可能性があると考えました。
まず、OCTN1が脳のどの細胞に存在するかを検討したところ、OCTN1はマウスの神経細胞に存在することがわかりました12)。一方で、グリア細胞には明確なOCTN1の存在は認められませんでした。培養した神経細胞ではERGOの細胞内への取り込みが確認された一方、octn1-/-から単離した神経細胞でERGOの取り込みは見られませんでした12)。したがって、OCTN1は神経細胞でERGOを取り込んでいることが分かりました。さらに研究を進めると、神経細胞以外にも、神経幹細胞13)とミクログリア14)にもOCTN1が存在することが分かりました。神経幹細胞は、自分自身が増殖して増える働き(自己増殖能)と神経細胞やグリア細胞へ変化する働き(多分化能)を持っています(図8)。
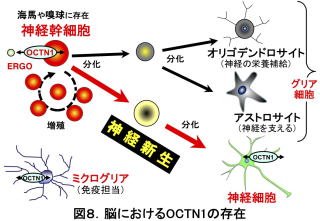
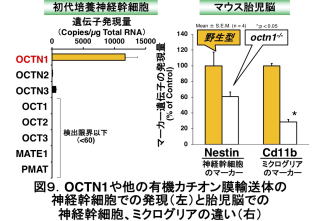
OCTN1の存在量は、同じ遺伝子familyに属する他の有機カチオン膜輸送体よりもはるかに高いものであり(図9)、神経幹細胞で何かの働きをしているかもしれません。そこで、マウス大脳皮質由来の培養神経幹細胞にERGOを添加したところ、神経細胞のマーカー(βIII
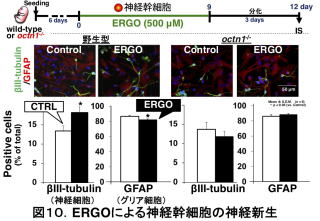
tubulin)の陽性細胞の割合が増加した一方、グリア細胞のマーカー(GFAP)の陽性細胞の割合が減少しました(図10)13)。つまり、神経幹細胞は成長すると神経細胞とグリア細胞の両方に変化しますが、ERGOはそのうちの神経細胞への分化(神経新生と呼びます)を促進することが示唆されます。octn1-/-から単離した神経幹細胞ではそのような作用は見えないため(図10)、ERGOの作用はOCTN1で取り込まれたことによって起こると言えます。ERGO以外の他の抗酸化物質(ビタミンCやエダラボンという薬物)をWTから単離した神経幹細胞に添加したところ、ERGOと同様に、抗酸化作用は発揮されました13)。しかし、ERGOで見られたような神経細胞への分化の促進作用は見られませんでした。このことは、ERGOによる神経新生の促進作用が、抗酸化作用とは異なるメカニズムによるものであることを示唆します。その後の解析から、神経幹細胞にERGOを添加すると、細胞内情報伝達経路の一種であるmTORC1(mammalian target of rapamycin complex 1)シグナルのタンパク質であるmTORと、その下流に位置するS6K1(p70 ribosomal protein S6 kinase)のリン酸化が増加するとともに、神経栄養因子neurotrophins (NT)の一種であるNT-5が増加することが分かりました15)。NT-5はその受容体であるTrkB(tropomyosin receptor kinase B)に結合することでTrkBを活性化し、脳のさまざまな働きを促します。ERGOの働きがNT-5という神経成長因子を介していることが示唆されます。なお、マウスの胎児の脳で観察すると、octn1-/-胎児脳では神経幹細胞やミクログリアのマーカー遺伝子の発現量(遺伝子がどのくらいONになったか)が低下していました(図9右)。したがって、OCTN1がおそらくERGOを取り込むことは、これら細胞自身の維持にも関わっていることが推測されます。
神経幹細胞は脳の海馬に多く存在し、アルツハイマー型認知症やうつ病患者では海馬の萎縮が起こることが知られています。また抗うつ薬の中には神経新生を促進するものがあります。このように神経新生は神経疾患と密接に関係します。そこで、ERGOの経口摂取による脳機能に及ぼす作用を調べました。マウスにERGOを多く含む餌を摂取させたところ、海馬の歯状回と呼ばれる部位にERGO濃度が顕著に増加し、未熟な神経細胞のマーカーであるdoublecortinの陽性細胞の数が増えることが分かりました16)。さらに、強制水泳試験や尾懸垂試験と呼ばれるうつ病モデルを用いたところ、ERGO摂取マウスでは、対照餌摂取群に比べ、自発運動量に変化がない一方、これら試験における無動時間が対照群と比べて短縮されました16)。ERGOの経口摂取に抗うつ作用のあることが示唆されました。
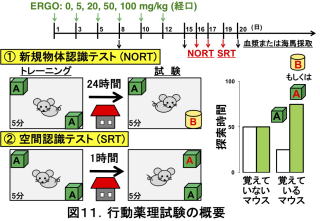
神経新生は神経の再生を意味します。したがって、脳の病態時だけでなく、正常な状態での脳機能にも影響を与える可能性があります。そこで、ERGO経口投与による記憶・学習能力に及ぼす影響を、通常飼育下の正常マウスに対して行いました。1日3回の経口投与を2週間行ったのち、novel
object recognition test (NORT)とspatial recognition test (SRT)と呼ばれる行動薬理試験を行い、それぞれ新規物体認識能と空間認識能へのERGO経口摂取の影響を調べました。これらの試験では、あらかじめ両隅に同じ物体を配置した立方体チャンバーにマウスを入れ、数分間のトレーニングを行い、マウスにこれら物体とその配置を覚えさせます(図11)。その後、一定時間飼育ケージに戻すことで記憶をほぼ消失させたのち、NORTにおいては片方の物体を別の物体に入れ、SRTにおいては片方をそのまま配置だけ変更させます(変化させた方をnovelと呼びます)。いずれにおいても、もう片方の物体は配置も含めそのままにしておきます(familiarと呼びます)。改めてマウスをチャンバーに入れて数分間の試験を行い、その間、novelな物体とfamiliarな物体を探索した時間を測定します。マウスは新しい物を探索する習性がありますので、トレーニング時の物体や配置を記憶していればnovelな物体に対する探索時間が延長し、覚えていなければ両方の物体に同程度の探索時間を示します(図11)。ERGOを経口投与後、NORTにおいて、novelな物体に対する探索時間は、familiarな物体への探索時間に比べて伸びました。SRTでも、NORTほどではありませんが同様な効果が見えました17)。この時のERGOの血漿中や脳海馬中濃度はERGOの投与量依存的に増加し、ERGOによる効果が裏付けられました。
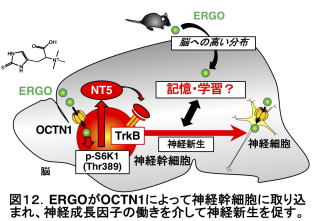
このような脳機能改善効果のメカニズムは未だによく分かっていません。ただ、最近の私たちの研究から、ERGOは脳の海馬という記憶学習に関わる領域で神経成長因子NT5というタンパク質を増やすこと、海馬に存在し、神経の再生に関わる神経幹細胞においてmTORC1シグナルと呼ばれる情報伝達経路を活性化することが分かってきました19)。NT5はさまざまな脳の働きを促進するタンパク質ですが、NT5をヒトやマウスに投与しても、血液脳関門と呼ばれるバリアが存在するためにNT5は脳にまで到達しません。しかし、ERGOであれば、脳の海馬でNT5を増やしてくれる訳ですから、NT5の作用を促すことができる訳です。
興味深いことは、このような脳機能促進効果が見られた血漿中ERGO濃度は、マウスやヒトの正常レベル(1 μM程度)の数倍以内であり、海外で実施された18)ヒトへのERGO経口投与試験(1日当たり 5ないし25 mg)後に見られる血漿中濃度とほぼ同じである点、すなわち、ヒトでも到達しうるレベルであることです。つまり、これまでの私たちの研究はマウスで行なってはいますが、そこで見られたERGOによる脳機能改善効果は、私たち人間でも起こり得て良い濃度で見られています。
そこでヒトでもERGOが脳に作用するのではないかと考え、ERGOのヒトにおける脳機能に及ぼす影響の解明を目的として、健常人と軽度認知障害(MCI, mild cognitive impairment)者を対象として、ERGOを含む食品を飲んでいただいた群と飲んでいただかなかった群(プラセボ群)とでさまざまな脳機能を比較する臨床研究を行いました。その結果、プラセボ群と比較して言語記憶の有意な増加が見えたほか、投与開始前に比べ投与後に総合記憶、言語記憶、認知機能速度、反応時間、持続的注意力が有意差に改善しました20)。つまり、ヒトにおいてもERGOが一部の脳機能を改善することが示唆されました。最近、ヒトにおけるERGOの血中濃度が、加齢や軽度認知障害、パーキンソン病などで減少することが相次いで報告されています21,22)。これらの結果は、脳機能や神経変性疾患においてもOCTN1ないしERGOが重要な働きを果たす可能性を示すと考えられます。食物から摂取したERGOが消化管から吸収されて脳の海馬に到達し、そこで神経成長因子の作用を介する形で神経新生を起こしながら、記憶学習能力を高めていると考えられます(図12)。
4.まとめ
膜輸送体の研究からスタートしたエルゴチオネイン物語は、ヒトがなぜ、自身が合成することのできないERGOのためにわざわざその膜輸送体を体内に用意しているのか?という問いからスタートしました。その答えの一つは、膜輸送体がないと細胞膜を通過できない水溶性アミノ酸であるERGOを体内に取り込み、炎症部位に作用させて炎症をコントロールするとともに、脳に作用させて脳機能を高めることかもしれません。つまりERGOは体内において生体を防御し脳機能を高めるビタミンのような働きをしていると考えられます。しかし現時点ではあくまでその状況証拠が掴めた段階です。今後さらに研究が発展することにより、ERGOが脳や他の組織においてビタミンとして働いていることが証明されるのではないかと考え、現在も研究を続けています。
【引用】
1) Tokuhiro S, Yamada R, Chang X, Suzuki A, Kochi Y, Sawada T, Suzuki M, Nagasaki M, Ohtsuki M, Ono M, Furukawa H, Nagashima M, Yoshino S, Mabuchi A, Sekine A, Saito S, Takahashi A, Tsunoda T, Nakamura Y, Yamamoto K. An intronic SNP in a RUNX1 binding site of SLC22A4, encoding an organic cation transporter, is associated with rheumatoid arthritis. Nat Genet 35(4): 341-348, 2003.
2) Peltekova VD, Wintle RF, Rubin LA, Amos CI, Huang Q, Gu X, Newman B, Van Oene M, Cescon D, Greenberg G, Griffiths AM, St George-Hyslop PH, Siminovitch KA. Functional variants of OCTN cation transporter genes are associated with Crohn disease. Nat Genet 36(5): 471-475, 2004.
3) Hou X, Mao J, Li Y, Li J, Wang W, Fan C, Wang H, Zhang H, Shan Z, Teng W. Association of single nucleotide polymorphism rs3792876 in SLC22A4 gene with autoimmune thyroid disease in a Chinese Han population. BMC Med Genet 16, 76, 2015.
4) Ben Said M, Grati M, Ishimoto T, Zou B, Chakchouk I, Ma Q, Yao Q, Hammami B, Yan D, Mittal R, Nakamichi N, Ghorbel A, Neng L, Tekin M, Shi XR, Kato Y, Masmoudi S, Lu Z, Hmani M, Liu X. A mutation in SLC22A4 encoding an organic cation transporter expressed in the cochlea strial endothelium causes human recessive non-syndromic hearing loss DFNB60. Human Genetics 135(5): 513-524, 2016.
5) Kato Y, Kubo Y, Iwata D, Kato S, Sudo T, Sugiura T, Kagaya T, Wakayama T, Hirayama A, Sugimoto M, Sugihara K, Kaneko S, Soga T, Asano M, Tomita M, Matsui T, Wada M, Tsuji A. Gene knockout and metabolome analysis of carnitine/organic cation transporter OCTN1. Pharm Res 27(5): 832-840, 2010.
6) Grundemann D, Harlfinger S, Golz S, Geerts A, Lazar A, Berkels R, Jung N, Rubbert A, Schomig E. Discovery of the ergothioneine transporter. Proc Natl Acad Sci USA 102(14): 5256-5261, 2005.
7) Sugiura T, Kato S, Shimizu T, Wakayama T, Nakamichi N, Kubo Y, Iwata D, Suzuki K, Soga T, Asano M, Iseki S, Tamai I, Tsuji A, Kato Y. Functional expression of carnitine/organic cation transporter OCTN1/SLC22A4 in mouse small intestine and liver. Drug Metab Dispos 38(10): 1665-1672, 2010.
8) Shimizu T, Masuo Y, Takahashi S, Nakamichi N, Kato Y. Organic cation transporter Octn1-mediated uptake of food-derived antioxidant ergothioneine into infiltrating macrophages during intestinal inflammation in mice. Drug Metab Pharmacokinet 30(3): 231-239, 2015.
9) Taubert D, Jung N, Goeser T, Schomig E. Increased ergothioneine tissue concentrations in carriers of the Crohn's disease risk-associated 503F variant of the organic cation transporter OCTN1. Gut 58(2): 312-314, 2009.
10) Tang Y, Masuo Y, Sakai Y, Wakayama T, Sugiura T, Harada R, Futatsugi A, Komura T, Nakamichi N, Sekiguchi H, Sutoh K, Usumi K, Iseki S, Kaneko S, Kato Y. Localizatoin of xenobiotic transporter OCTN1/SLC22A4 in hepatic stellate cells and its protective role in liver fibrosis. J Pharm Sci 105(5): 1779-1789, 2016.
11) Shinozaki Y, Furuichi K, Toyama T, Kitajima S, Hara A, Iwata Y, Sakai N, Shimizu M, Kaneko S, Isozumi N, Nagamori S, Kanai Y, Sugiura T, Kato Y, Wada T. Impairment of the carnitine/organic cation transporter 1-ergothioneine axis is mediated by intestinal transporter dysfunction in chronic kidney disease. Kid Int 92(6): 1356-1369, 2017.
12) Nakamichi N, Taguchi T, Hosotani H, Wakayama T, Shimizu T, Sugiura T, Iseki S, Kato Y. Functional expression of carnitine/organic cation transporter OCTN1 in mouse brain neurons: Possible involvement in neuronal differentiation. Neurochem Int 61(7): 1121-1132, 2012.
13) Ishimoto T, Nakamichi N, Hosotani H, Masuo Y, Sugiura T, Kato Y. Organic cation transporter-mediated ergothioneine uptake in mouse neural progenitor cells suppresses proliferation and promotes differentiation into neurons. PLoS One 9(2): e89434, 2014.
14) Ishimoto T, Nakamichi N, Nishijima H, Masuo Y, Kato Y. Carnitine/organic cation transporter OCTN1 negatively regulates activation in murine cultured microglial cells. Neurochem Res 43(1): 107-119, 2018.
15) Ishimoto T, Masuo Y, Kato Y, and Nakamichi N. Ergothioneine-induced neuronal differentiation is mediated through activation of S6K1 and neurotrophin 4/5-TrkB signaling in murine neural stem cells. Cell Signal 53: 269-280, 2019.
16) Nakamichi N, Nakayama K, Ishimoto T, Masuo Y, Wakayama T, Sekiguchi H, Sutoh K, Usumi K, Iseki S, Kato Y. Food-derived hydrophilic antioxidant ergothioneine is distributed to the brain and exerts antidepressant effect in mice. Brain Behav 6(6): e00477, 2016.
17) Nakamichi N, Nakao S, Nishiyama M, Takeda Y, Ishimoto T, Masuo Y, Matsumoto A, Suzuki M, and Kato Y. Oral administration of the food derived hydrophilic antioxidant ergothioneine enhances object recognition memory in mice. Curr Mol Pharmacol, in press.
18) Cheah IK, Tang RM, Yew TS, Lim KH, Halliwell B. Administration of Pure Ergothioneine to Healthy Human Subjects: Uptake, Metabolism, and Effects on Biomarkers of Oxidative Damage and Inflammation. Antioxid Redox Signal 26(5): 193-206, 2017.
19) Ishimoto T, Masuo Y, Kato Y, and Nakamichi N. Ergothioneine-induced
neuronal differentiation is mediated through activation of S6K1 and neurotrophin
4/5-TrkB signaling in murine neural stem cells. Cell Signal 53: 269-280,
2019.
20) 渡邉憲和、松本 聡、鈴木 真、深谷泰亮、加藤将夫、橋弥尚孝 健常者および軽度認知障害者に対するエルゴチオネイン含有食品の認知機能改善効果
ランダム化プラセボ対照二重盲検並行群間比較試験、薬理と治療 48(4): 685-697, 2020.
21) Cheah IK, Feng L, Tang RMY, Lim KHC and Halliwell B. Ergothioneine
levels in an elderly population decrease with age and incidence of cognitive
decline; a risk factor for neurodegeneration? Biochem Biophys Res Commun
478(1): 162-167, 2016.
22) Hatano T, Saiki S, Okuzumi A, Mohney RP and Hattori N. Identification
of novel biomarkers for Parkinson's disease by metabolomic technologies.
J Neurol Neurosurg Psychiatry 87(3): 295-301, 2016.
① 薬物治療を評価するバイオマーカーの探索研究
詳細は、こちら。
② 抗がん薬等の最適な薬物治療を目指した研究
詳細は、こちら。
③ 健康食品、ビタミンの探索による病気予防の研究
詳細は、こちら。
④ 生理活性タンパク質の体内動態とアンメット疾患への応用
詳細は、こちら。
分子薬物治療学研究室ビルダークリニック
〒920-1192
石川県金沢市角間町
金沢大学薬学系
bunyaku@p.kanazawa-u.ac.jp

![]()
![]()
